+Neural plasticity in adult somatic sensory motor system pdf
Teks penuh
Gambar
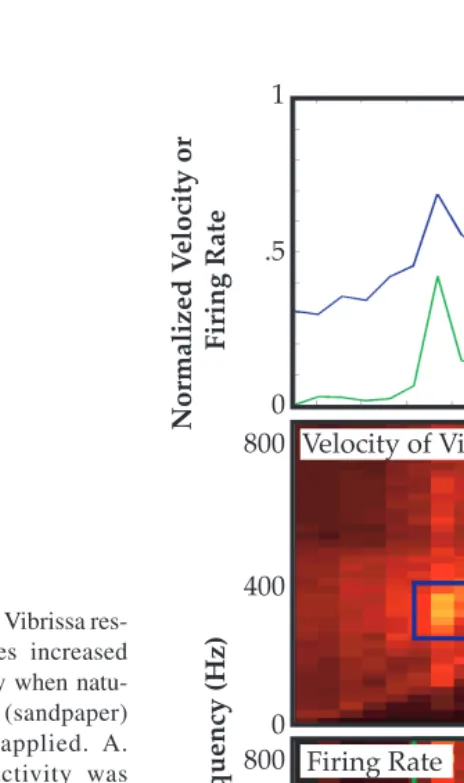
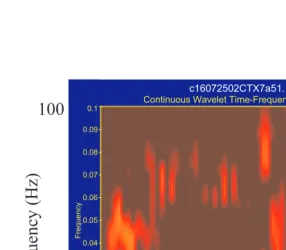
![FIGURE 2.10] An example of the temporal evolution of neural frequency tuning. A. TopPeri-stimulus time histograms (PSTHs) are plotted as a function of frequency of stimulation(ordinate) and time (abscissa)](https://thumb-ap.123doks.com/thumbv2/123dok/3941703.1885123/26.612.63.384.76.388/temporal-evolution-frequency-histograms-function-frequency-stimulation-abscissa.webp)

Dokumen terkait
SEGMEN BERITA REPORTER A Sry Tuntut Para Pengusaha Yang Mengabaikan
Sistem ini hanya sebuah media pencarian rumus-rumus serta simulasi soal-soal matematika tingkat SLTA untuk membantu para siswa menjawab soal matematika serta
Dengan ini saya menyatakan skripsi yang berjudul “ Profil Kejenuhan Belajar Siswa Berdasarkan Latar Belakang Biografis Serta Implikasinya Bagi Bimbingan dan Konseling
Evaluasi dan indikator keberhasilan intervensi solution focused briefcounseling dalam setting kelompok untuk mengingkatkan self-control siswa pada sesi 3, sebagai
Tujuan penelitian adalah untuk membandingkan plasma protein darah antara pre-albumin ( Pa ) dan albumin ( Alb ) pada domba beranak tunggal (D1), beranak kembar dua (D2) dan
penelitian ini adalah mengetahui : (1) pertumbuhan kapulaga di bawah keempat jenis tegakan pohon dalam pola tanam agroforestry dan (2) karakteristik tanah di
Character Building: Optimalisasi Peran Pendidikan dalam Pengembangan Ilmu dan Pembentukan Karakter Bangsa.. Jogjakarta: Ar-Ruzz
11 Pianta and others, “Effects of Web-Mediated Profes- sional Development Resources on Teacher-Child Interactions in Pre-Kindergarten Classrooms ”; Allen and others,
