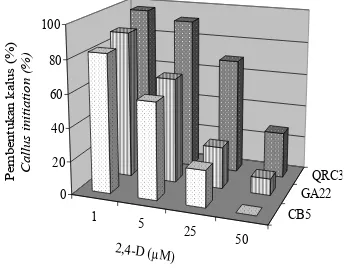
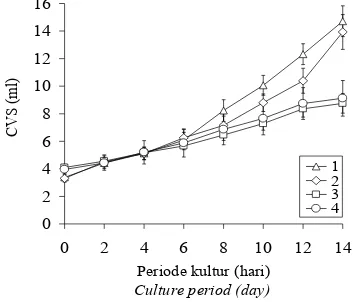
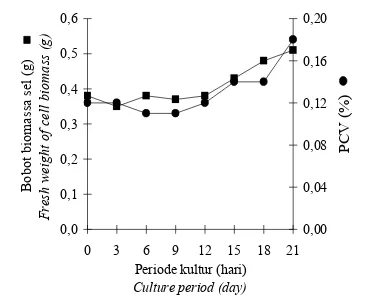
Pertumbuhan biak kalus dan suspensi sel tanaman kina (Cinchona ledgeriana Moens)
Teks penuh
Gambar
Dokumen terkait
Menurut status kelembagaan lahan, produktivitas lahan petani pemilik penggarap lebih tinggi dibandingkan dengan penyewa sedangkan produktivitas lahan penyewa lebih tinggi
Hasil uji statistik Chi-square, diperoleh nilai p = 0,332 yang berarti p > 0,05 sehingga dapat disimpulkan bahwa tidak terdapat hubungan signifikan antara keberadaan
Di Indonesia sendiri, sebuah penelitian di Denpasar dengan subjek anak-anak menunjukkan bahwa pada awal infeksi virus dengue, penderita yang mengalami
Sekali lagi perlu dipahami bahwa sasaran atau objek sebagaimana dimaksudkan di atas adalah sesuatu yang bersifat nyata atau yang senyatanya ada ( das sein ), bukan
Uji homogenitas dilakukan melalui pengujian terhadap hasil pretest dan posttest pada kemampuan pemecahan masalah ranah kognitif dari kedua kelas, uji homogenitas
“ Selain itu, transaksi dalam keuangan Selain itu, transaksi dalam keuangan Islam sesuai dengan syariah harus. Islam sesuai dengan
bahwa untuk melaksanakan ketentuan Pasal 2 ayat (6) Peraturan Menteri Hukum dan Hak Asasi Manusia Nomor 26 Tahun 2020 tentang Visa dan lzin Tinggal dalam Masa Adaptasi
Berdasarkan dari analisis data yang diperoleh, dapat ditarik kesimpulan diantaranya (1) kemampuan bercerita siswa yang diberikan permainan dramatik lebih tinggi daripada siswa
