Keragaman Genetik DNA Mikrosatelit dan Hubungannya dengan Performa Bobot Badan pada Domba Lokal
Teks penuh
Gambar
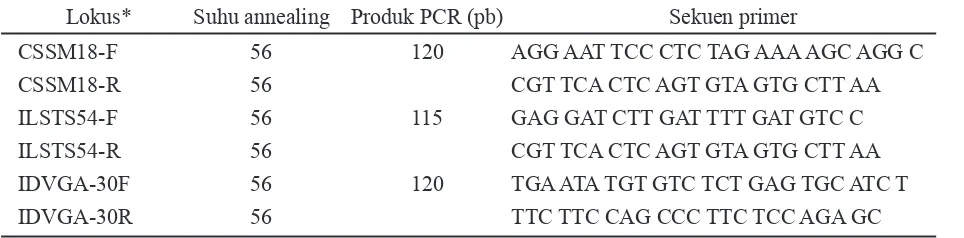
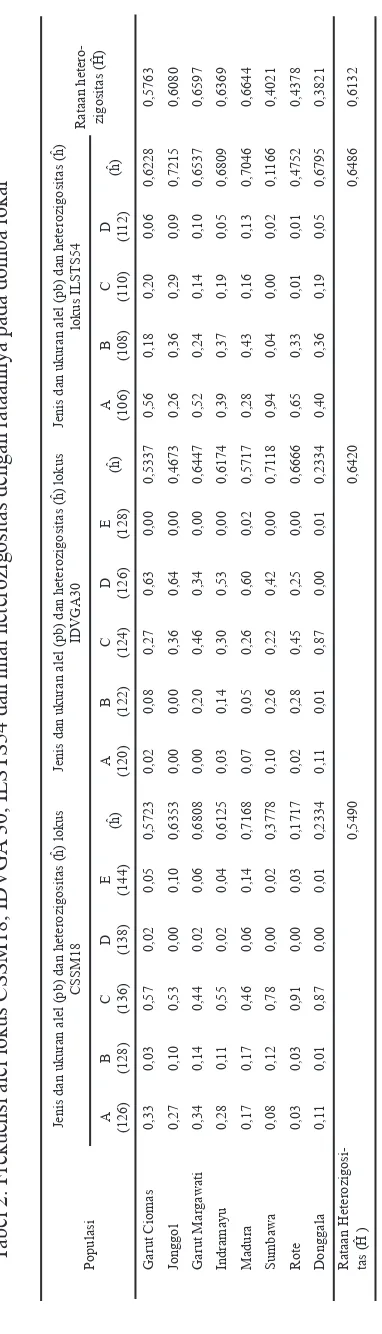
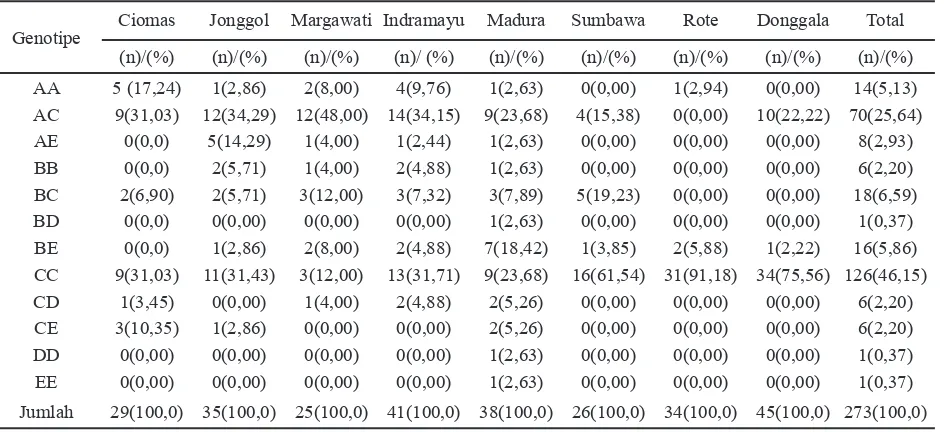
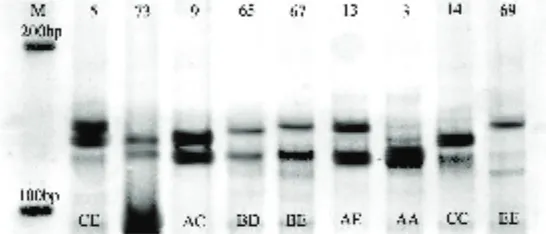
Dokumen terkait
Dengan dilakukannya observasi ini diharapkan mahasiswa lebih bisa mempersiapkan pembelajaran yang akan dilakukan dari segi metode, pengelolaan kelas dan lain
Kota Pangkalpinang dengan luas wilayah paling kecil memiliki aliran permukaan yang kecil, sedangkan aliran permukaan terbesar terdapat pada Kabupaten Bangka
Dalam tugas akhir ini akan direncanakan struktur jembatan menggunakan busur rangka batang baja yang melewati sungai Grindulu, Kabupaten Pacitan dengan bentang total 354
Dari novel tersebut seakan membawa kita pada suatu makna bahwa jika kita memiliki kesukaan terhadap sesuatu yang benar-kita sukai selagi itu merupakan hal yang positif
Hasil analisis regresi linear menunjukkan hubungan yang signifikan secara statistik antara kepercayaan diri dengan prestasi belajar dengan p =.0,015 Sedangkan
Sistem ini hanya akan melakukan Pengolahan terhadap data pendaftaran siswa baru dan registrasi siswa per satu semester ajaran akademik yang aktif tercatat sebagai
Apabila jumlah aset yang dimiliki perusahaan meningkat maka informasi yang diungkapkan perusahaan akan semakin banyak dan lengkap sehingga mampu meyakinkan investor
Perbedaan dari hasil penelitian terdahulu dengan penelitian yang saya teliti sekarang yaitu, penelitian yang saya lakukan sekarang lebih ke banyaknya nasabah yang
