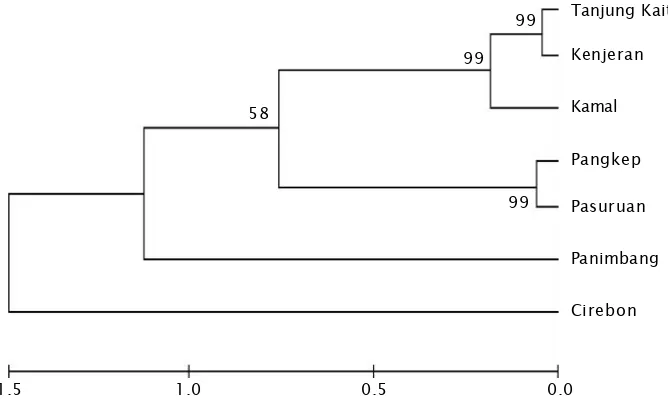
HUBUNGAN KEKERABATAN BEBERAPA POPULASI KERANG HIJAU (Perna viridis) DI INDONESIA BERDASARKAN SEKUEN CYTROCROME B mtDNA
Teks penuh
Gambar
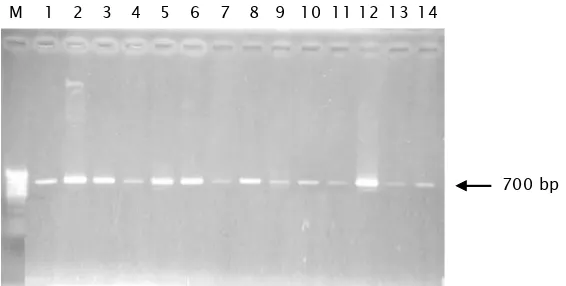
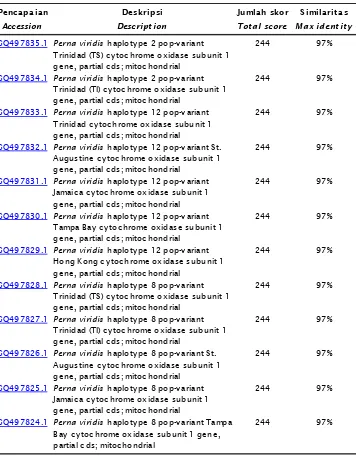
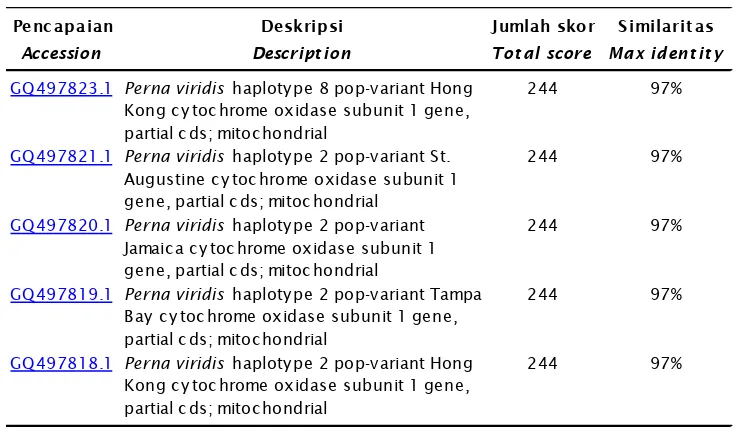
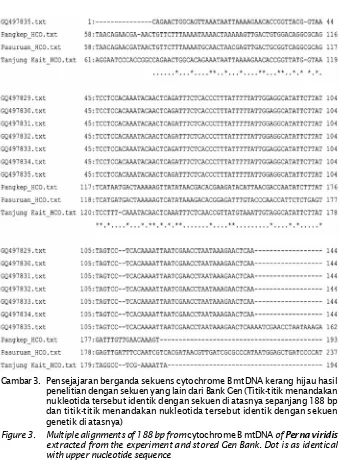
Dokumen terkait
Pengaruh human relations terhadap kepuasan kerja karyawan pada bagian logistik Di pt badjatex bandung!. Universitas Pendidikan Indonesia | repository.upi.edu
50:50 sesuai dengan hasil penelitian Gurav and Pathade (2011) pada penelitian pembuatan vermikomposting dengan media pertumbuhan cacing tanah menggunakan capuran kotoran
Risdawati Lubis : Uji Jarak Cerobong Udara Dan Lama Pengeringan Terhadap Mutu Kunyit Kering Alat Pengering...,2005.. un JAKAK CEROBONG UDAKA D4.N LAMA PENGERINGAN TERHADAP MUTU
Keadaan gizi buruk pada anak yang banyak terjadi di Provinsi Jambi salah satunya adalah kwashiorkor, yaitu suatu kondisi seseorang yang kekurangan protein dibawah
(1) Orang tua atau wali dari Pecandu Narkotika yang belum cukup umur wajib melaporkan kepada pusat kesehatan masyarakat, rumah sakit, dan/atau lembaga
Judul Skripsi : PENGARUH INTENSITAS MEMBACA ASMAUL HUSNA PADA AWAL PEMBELAJARAN TERHADAP KONSENTRASI SISWA DALAM MENGIKUTI PEMBELAJARAN ( Studi Pada siswa
Kemudian dari data gas lift valve yang ada di- input ke dalam PROSPER untuk dianalisis, sehingga kita dapat membuat model existing gas lift design yang ada saat
Dalam hal daerah telah mempunyai Peraturan Daerah sebagaimana dimaksud pada ayat (1) sebelum Peraturan ini diberlakukan, maka Peraturan Daerah tersebut harus
