RESPON INTERFERON GAMMA TERHADAP Plasmodium falciparum RADIASI PADA KULTUR SEL LIMFOSIT MANUSIA - e-Repository BATAN
Teks penuh
Gambar
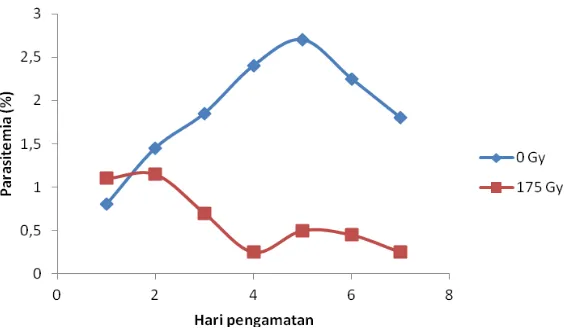
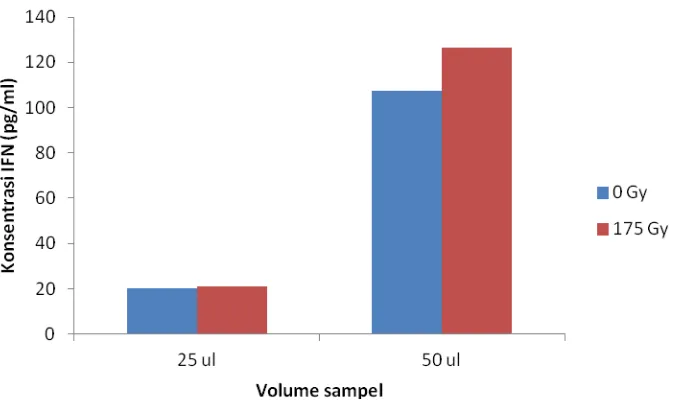
Dokumen terkait
Aqidah tidak boleh hanya dipahami sebagai keyakinan pada Rukun Iman saja, yaitu iman pada Allah, malaikat Allah, Kitab-kitab Allah, nabi, hari akhir, dan
Puji syukur penulis panjatkan kehadirat Allah SWT, karena atas berkat dan rahmat-Nya yang telah memberikan kemudahan dalam menyelesaikan skripsi ini yang berjudul Representasi
Dari analisis yang telah didapat berdasarkan perhitungan menggunakan metode OCRA Index, kemudian akan diperoleh hasil berupa usulan perbaikan alat kerja
Ukuran dari kateter intravena juga mempengaruhi kejadian phlebitis, pemasangan kateter intravena yang berukuran besar (bernomor kecil) pada vena yang kecil akan menyebabkan
adalah terlalu perlahan.. Data BOD ialah ujian utama untuk menilai kepekatan atau kekuatan air buangan domestik, industri dan pertanian dari segi kandungan oksigen
Intan Pariwara semakin membuktikan signifikansi teori dan juga para peneliti terdahulu yang digunakan sebagai referensi dalam penelitian ini, khususnya dari segi
Hasil analisis kekerasan kedua kelongsong bahan bakar tersebut dapat dinyatakan bahwa paduan AlMgSi lebih kuat untuk digunakan sebagai kelongsong bahan bakar U 3 Si
Sesuai dengan penelitian yang dilakukan Syahrani (2006) substansi CO merupakan hasil gabungan karbon dan oksigen, dimana gabungan tersebut tidak mencukupi untuk
