Pengaruh Kombinasi NAA Dengan Sitokinin (BAP, Kinetin dan 2iP) Terhadap Daya Ploriferasi Tanaman Kantong Semar (Nepenthes mirabilis) Secara In Vitro.
Teks penuh
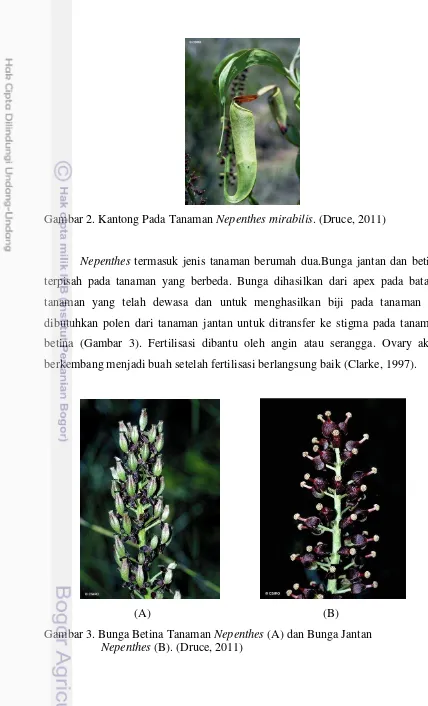
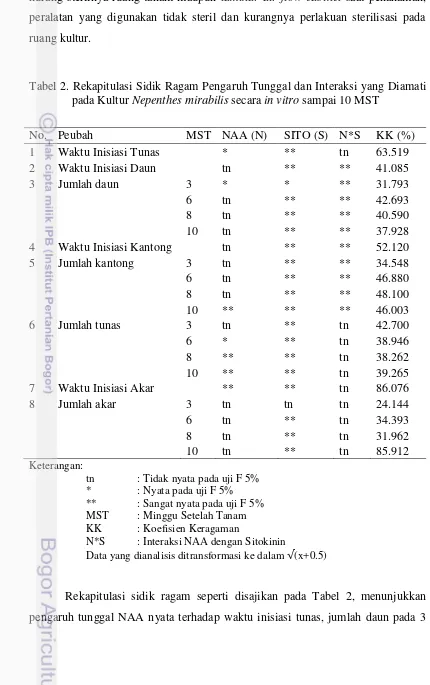
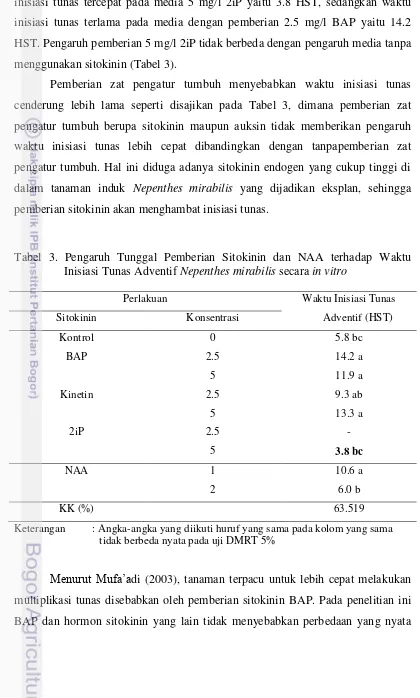
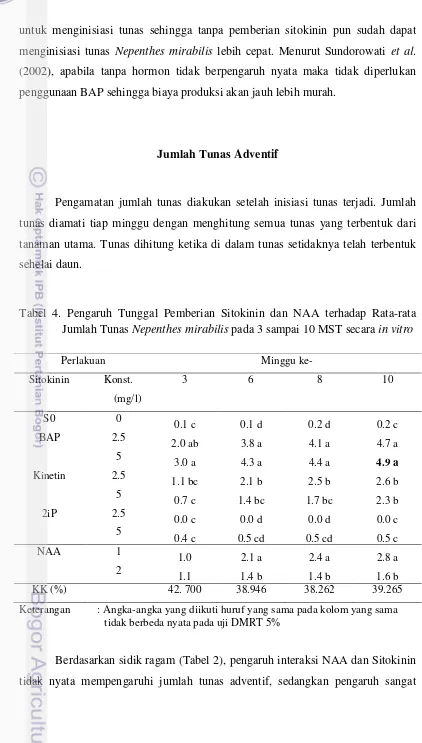
Gambar


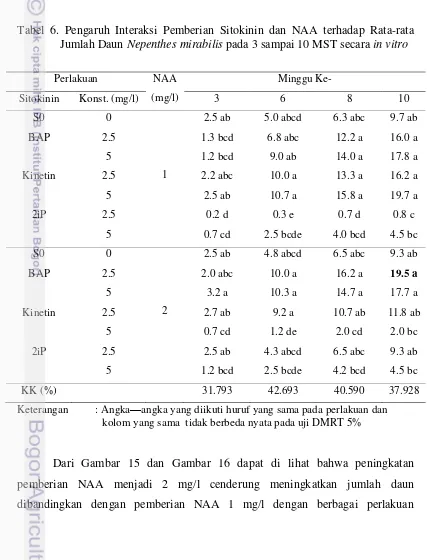
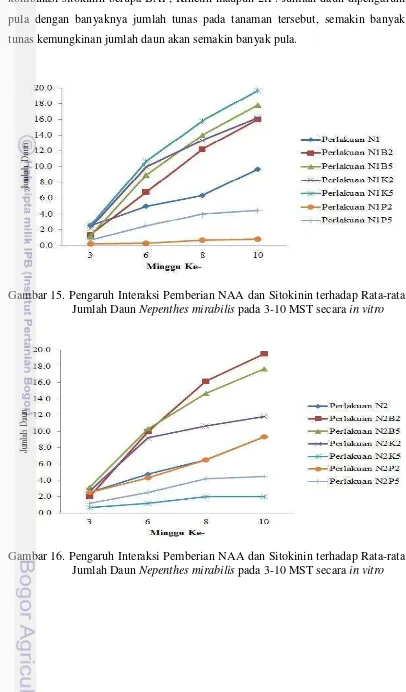


Dokumen terkait
perbedaan hasil belajar siswa dengan menggunakan model pembelajaran kooperatif tipe Numbered Head Together (NHT) dan Two Stay Two Stray (TSTS) pada materi sistem
Cara lain untuk mengatasi IGSCC adalah dengan pemberian panas pada material yang rentan terhadap IGSCC, cara ini telah diterapkan pada reaktor Fugen yang dikenal. dengan metoda
Sehubungan dengan hal tersebut, penulis membuat aplikasi pembelajaran bahasa Inggris disertai kamus dengan menggunakan metode aplikasi yang berbasis web yang bersifat
Penelitian ini berhasil menyusun rancangan operational information system dengan mengintegrasikan keseluruhan data yang ada di Kepolisian, Pengadilan Negeri, Kejaksaan
Berdasarkan Penetapan Pemenang Pelelangan Nomor : 98/ULP-POKJA.JL/III/2016 Tanggal 30 Maret 2016 dan Berita Acara Hasil Pelelangan Nomor : 95/ULP-POKJA.JL/III/2016 Tanggal
Penelitian dalam tesis ini dilatarbelakangi oleh munculnya perjanjian jual beli online yang timbul sebagai suatu kendala tentang perjanjian, perpajakan, tata cara
Selanjutnya preskripsi pengaturan hasil pada hutan bekas tebangan lain yang tidak memiliki PUP, dengan menggunakan model komponen dinamika struktur tegakan yang diperoleh
Pulau Sakanun merupakan pulau sangat kecil, memiliki terumbu karang fringing reef, ekosistem lamun berada di sekitar pesisir pulau dan kawasan intertidal yang
