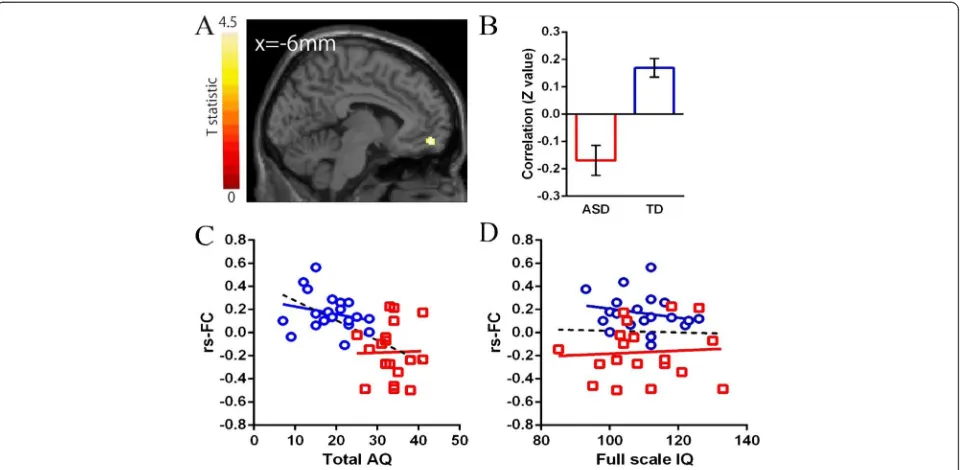
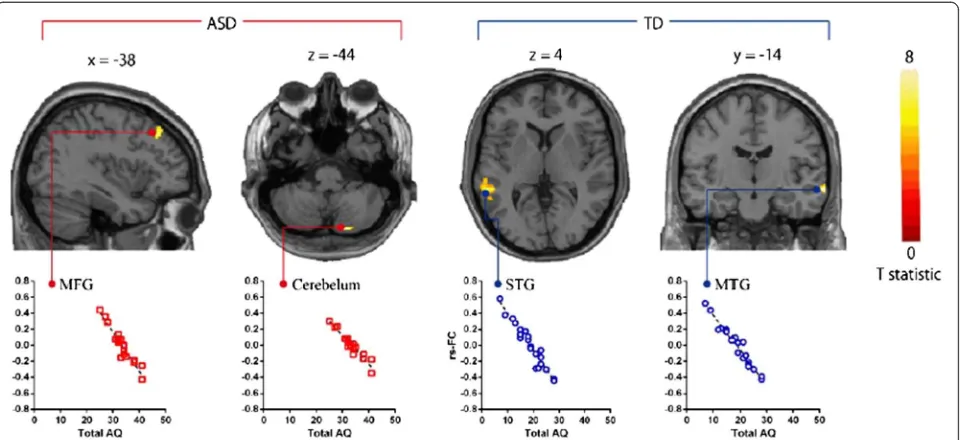
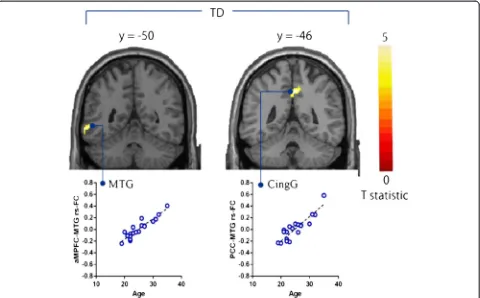
Default mode network in young male adult
Teks penuh
Gambar
Garis besar
Dokumen terkait
Meskipun dalam Pasal 24 UUD 1945 tidak menyebutkan secara tersurat mengenai kewenangan Mahkamah Agung dalam proses seleksi dan pengangkatan calon hakim dari lingkungan
Berdasarkan surat penetapan penyediaan barang dan jasa Nomor 03/PPBJ.04.02/III/2015 tanggal 27 Maret 2015, dengan ini pejabat pengadaan barang dan jasa Dinas pertanian
Hasil Total Analisis Kadar Protein dan Lemak Ikan Julung-julung Asap ( Hemiramphus far ). No Sampel Kadar Protein
Ekstrak etanol umbi ubi jalar ungu ( Ipomoea batatas L.) mengandung flavonoid jenis antosianin yang memiliki aktivitas sitotoksik dan menghambat proliferasi.
Tes Tertulis : melakukan penjumlahan dan pengurangan, perkalian dan pembagian bilangan asli , bilangan bulat dan pecahan sesuai prosedur/aturan dengan memperhatikan nilai
Judul Skripsi : PENGARUH STRUKTUR DAN MEKANISME CORPORATE GOVERNANCE TERHADAP TINGKAT KEPATUHAN MANDATORY DISCLOSURE (Studi Empiris Pada Perusahaan Manufaktur yang
SPK merupakan sistem yang sangat membantu lurah dalam penentuan keluarga miskin dalam mendapatkan bantuan, SPK yang digunakan dengan metode AHP.. AHP merupakan sistem yang
PENGARUH PEMBELAJARAN PERMAINAN HOKI TERHADAP KEBUGARAN JASMANI DAN KEPERCAYAAN DIRI SISWA DI SMA NEGERI 26 GARUT.. Universitas Pendidikan Indonesia | repository.upi.edu
