MATERIALS AND METHODS Materials
Teks penuh
Gambar
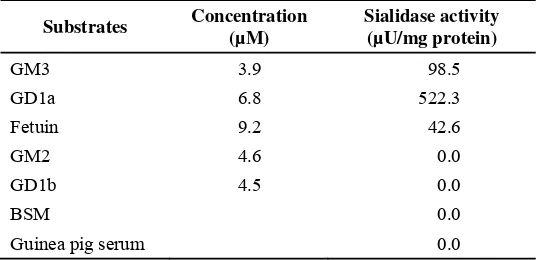
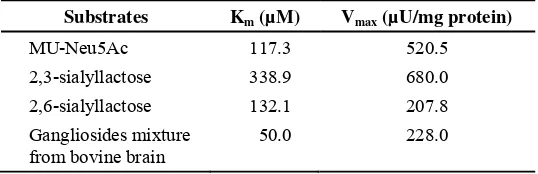
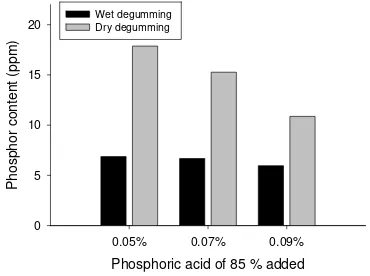
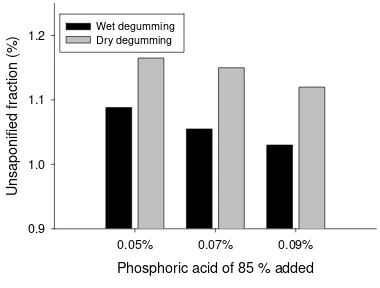
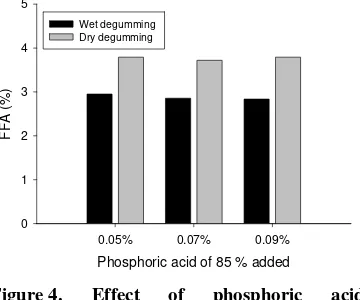
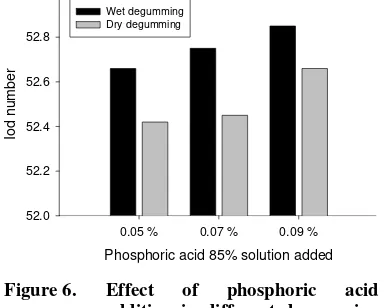
Garis besar
Dokumen terkait
Untuk mendapatkan data yang terjadi pada fenomena sosial yang ada dalam lapangan, banyak jenis dan sumber data yang dapat digunakan tetapi tidak semua teknik itu
Manajemen perusahaan merupakan pihak yang menerapkan fungsi-fungsi yang meliputi perencanaan, pengorganisasian, pemotivasian, pengelolaan staf dan pengendalian. PT Kuala Pangan
Dengan hasil pengujian yang dilakukan terhadap variabel kualitas pelayanan (X1), menjelaskan bahwa kualitas pelayanan memiliki pengaruh yang signifikan
Mahasiswa yang berperan sebagai komunikator dalam ruang peradilan semu, sebagian informan yang mengalami ketakutan berkomunikasi dapat menyebabkan sebuah diskusi kurang
ditambahkan untuk setiap stoples kaca dan ditambah juga SCOBY dengan berat sesuai dengan perlakuan (5%, 10%, 15%) ke dalam setiap stoples kaca yang telah terisi teh bunga
Informan juga menuturkan meskipun boto putih merupakan pemakaman khusus bagi para keturunan raja, yang datang ke tempat tersebut berasal dari semua golongan, baik
Dari hasil FGD dan wawancara kepada mereka diperoleh gambaran bahwa meski memiliki lahan yang subur, namun tata produksi pertanian mereka belumlah seoptimal petani lain
Tujuan penelitian adalah untuk mengetahui pengaruh pemupukan nitrogen jangka panjang dengan berbagai dosis terhadap pertumbuhan dan produksi padi gogo, mengetahui pengaruh
