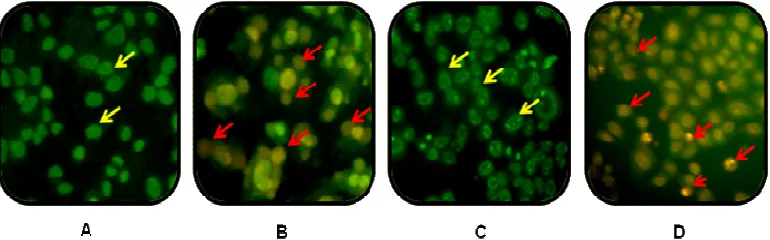
Hesperidin meningkatkan efek sitotoksik doxorubicin pada sel MCF-7
Teks penuh
Gambar
Dokumen terkait
Berdasarkan literatur tentang risiko fiskal yang dikembangkan oleh Brixi (2001) dan Cottarelli (2014), dan telaah dokumen Nota Keuangan dan Rancangan Anggaran
Tujuan penelitian ini untuk mengetahui dan mengevaluasi sistem penerimaan Biaya Penyelenggaraan Ibadah Haji (BPIH) pada Bank Syariah dan Konvensional yang telah
Tanah yang ada di Sidoresmo Dalam diberikan oleh pemerintah kolonial Belanda kepada Kiai Mas Cholil dan kawan-kawansebagai tanah perdikan yang bebas dari pajak
Data yang penulis dapatkan selama melakukan kegiatan Kuliah Pengabdian Masyarakat (KPM) adalah dengan melakukan pengamatan langsung di Desa Sepat dan
Hasil analisis distribusi untuk alasan mahasiswa memilih tempat mengerjakan tugas kuliah individu menunjukan bahwa kategori kenyamanan (muncul sebanyak 52 kali)
Ketidaksesuaian tersebut antara lain dalam hal: struktur tim manajemen yang hanya terdiri dari penanggungjawab dan bendahara, tidak memiliki anggota dari unsur orangtua siswa di
Menimbang bahwa setelah Majelis Hakim Tingkat Banding membaca Salinan Putusan Pengadilan Agama Bekasi a quo, memori banding dan kontra memori banding dan mencermati Berita
Kriteria ini mengukur sejauh mana sistem informasi dapat mendukung strategi perusahaan dalam meningkatkan kapasitas kerja dari pegawai dalam melakukan tugas dan fungsinya
