Synbiotic Application for Pacific White Shrimp Litopenaeus vannamei Resistance to Infectious Myonecrosis Virus and Growth
Teks penuh
Gambar





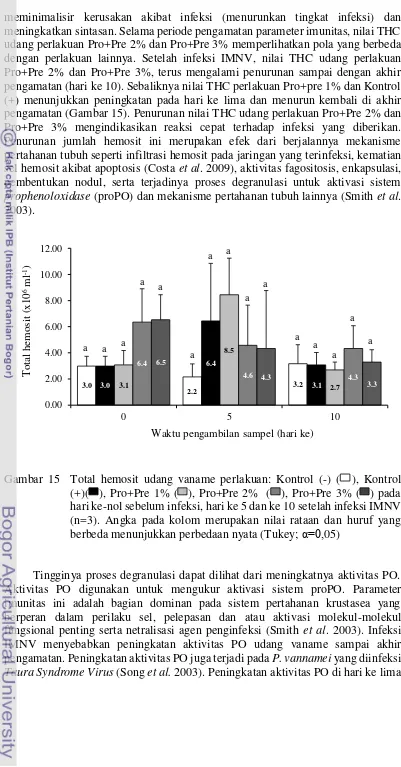
Dokumen terkait
Sementara itu, defisit perdagangan AS dengan Tiongkok juga turun menjadi US$37,9 miliar pada November 2018 dari US$43,1 miliar pada Oktober 2018.. Pencapaian ini melebihi target
Dalam al-Qur’an membahas masalah ini yang mana uraian ini dimulai dengan sejarah Israil, beranjak kesejarah orang-orang Kristen dan akhirnya kepada orang-orang Islam. dinyatakan
Kendaraan Bis Antar Kota Dalam
Selanjutnya adalah diskusi kelompok ahli, dalam diskusi kelompok ahli mereka lebih efektif mendengar penjelasan dari siswa kategori tinggi dan mendengar pertanyan dari
Berdasarkan hasil analisis, terdapat potensi dan permasalahan dalam menanggapi daya tarik lokasi wisata Pantai Tablanusu, hal yang dinikmati para pengunjung yang datang,
Nilai rata-rata Responden Ahli pada tahap ketiga adalah 1,81 dan nilai mediannya yaitu 2,00, artinya bahwa sub faktor adanya industri pendukung dan
7.. 4) Adanya perubahan organisasi, down sizing dan relokasi karyawan secara perlahan berpengaruh pada pencapaian usaha PPI. Kerugian demi kerugian terus menerus
tentang sesuatu pada saat yang timbul secara tiba-tiba, sedangkan kata belum mengandung ciri makna proses, peristiwa atau keadaan yang melibatkan jangka waktu
